Gestion non chirurgicale des déviations angulaires chez le poulain
Résumé
Les aplombs du cheval correspondent à l’orientation des membres sous le corps. Le jeune poulain peut être confronté quelques jours après sa naissance ou durant sa croissance à des déviations angulaires de ces membres entrainant une mauvaise répartition des charges, une usure non homogène au niveau des articulations, des muscles et des ligaments. Ces déviations sont à l’origine de pathologies telles que l’ostéochondrose, de l’ostéo-arthrose, des tendinites ou des fractures, susceptibles d’influencer ses performances et sa carrière par la suite. Ces déviations angulaires concernent principalement les articulations du carpe, du boulet et du jarret. Lors des déviations congénitales, elles sont attribuées à l'immaturité de la structure osseuse, à une laxité des structures péri-articulaires, une croissance osseuse asynchrone ou à une déviation diaphysaire. Dans le cas des déviations acquises, la croissance osseuse disproportionnée des plaques de croissance et des centres d’ossification des épiphyses en sont les responsables. Le traitement de ces déviations, doit être précoce avant la fermeture des plaques de croissance du poulain. Divers traitements sont actuellement disponibles pour corriger ces déviations, il s’agit des mesures conservatrices, de la maréchalerie ou des interventions chirurgicales. Le but de cette mise à jour bibliographique est de recenser les différents traitements non chirurgicaux décris actuellement et de comparer leurs résultats, afin de garantir aux vétérinaires praticiens équins et aux étudiants vétérinaires la meilleure prise en charge thérapeutique de la pathologie.
Mot clés: déviations angulaires, traitement, valgus, varus, poulain
Téléchargements
INTRODUCTION
Les déviations angulaires sont définies comme une déformation posturale axiale du membre vu dans le plan frontal (Smith, 2010; Guillot et Lallemand, 2019). Elles sont généralement associées à un certain degré de rotation interne ou externe (Auer, 1999). Chaque déviation est décrite selon la direction de la déformation, l’articulation touchée et le degré de déviation. Ainsi, on distingue deux types de déviations angulaires (Witte et Hunt, 2009; Guillot et Lallemand, 2019):
• Le varus est une déviation médiale du membre, distalement à la zone touchée (Figure 1-A);
• Le valgus est une déviation latérale du membre, distalement à la zone atteinte (Figure 1-B).
Le degré de la déviation varie d’une déviation discrète (<5°), modérée (5-10°), marquée (10-15°) à sévère (>15°). La graduation ne préjuge pas la gravité de l’affection. En effet, la gravité dépend de l’articulation atteinte, du type de déformation et de l’âge du poulain (Betsch, 2005; Guillot et Lallemand, 2019).
La déviation angulaire peut toucher des structures variées (Watson, 2016; Guillot et Lallemand, 2019):
• Le cartilage de croissance ou physe d’un os long: C’est le type de déviation le plus commun. Les atteintes les plus fréquentes sont celles de la physe distale du radius et de la physe distale de l’os métacarpien/métatarsien III (Figure 2-A);
• Une articulation: Le valgus du carpe est la déviation articulaire la plus fréquente, mais des déviations du jarret sont également assez courantes (Figure 2-B);
• La diaphyse d’un os long: Ces déformations sont les plus rares. Elles nécessitent une évaluation et une stratégie de traitement différentes de celles des malformations précédentes (Figure 2-C).
ÉTIOLOGIE
Les déformations angulaires des membres peuvent être classées dans deux grandes catégories d’étiologies différentes (Witte et Hunt 2009; Robert et al., 2013; Smith, 2015; Dubois et Rodgerson, 2020):
• Les déviations congénitales: elles sont présentes à la naissance ou peu après. Elles sont attribuées soit à l’immaturité de la structure osseuse (défaut d’ossification ou ossification incomplète (hypoplasie) des os (Figure 3A), soit à une laxité des structures péri-articulaires, une croissance osseuse asynchrone (Figure 3B), ou à une déviation diaphysaire (Figure 3C). Les principaux facteurs de cette déformation congénitale sont liés soit à la gestation (gestation gémellaire, placentite, prématurité, mauvaise position inta-utérine), soit à des facteurs maternels (nutrition, parasitisme, hypothyroïdisme) (Morisset, 2005).
• Les déviations acquises sont le résultat d’une croissance osseuse disproportionnée des plaques de croissances (des charges dissymétriques trop importantes le long de la plaque de croissance) et des centres d’ossification des épiphyses dans les premières semaines à quelques mois de la vie du poulain, avec plusieurs facteurs postnataux mis en cause: mauvaise conformation, alimentation déséquilibrée, exercice excessive, traumatisme externe, ostéomyélite (Morisset, 2005; Witte et Hunt, 2009; Bischofberger et Auer, 2019).
DIAGNOCTIC
Le diagnostic des déviations angulaires est basé sur l’examen clinique et radiographique. Il devra identifier, le nombre, la nature, l’angle et la localisation de la charnière de chaque déviation. Le diagnostic permettra, entre-autres, l’identification de la cause sous-jacente de la déviation, qui n’est pas toujours évidente, et conditionnera la conduite à tenir et le traitement à apporter (Trumble, 2005; Bramlage et Auer, 2006; Wilson, 2006; Ernst et al., 2020).
Diagnostic clinique
Le diagnostic clinique doit être basé sur un examen statique et dynamique complet afin d’évaluer le poulain dans son ensemble. Chaque membre doit être examiné de face, de derrière, de profil en appréciant avec soin sa direction et sa morphologie. Par la suite, le pied peut être soulevé, en saisissant l’os du canon parallèle au radius ou au tibia, tout en fléchissant doucement le carpe ou le tarse. Ceci dans le but d’évaluer tout écart par rapport à l’alignement axial et de différencier entre des déformations complexes impliquant plusieurs articulations (Figure 4) (Auer, 2006; Bramlage et Auer, 2006; Smith, 2010; Redden, 2011). Au même temps, le membre est palpé dans l’intention d’apprécier si la déviation est réductible manuellement, partiellement améliorée ou impossible à modifier, de déceler la présence ou non de douleur, d’enflure, d’épanchements synoviaux, d’une instabilité latéro-médiale ou d’une instabilité cranio-caudale (Greet, 2000; Betsch, 2005; Trumble, 2005; Bramlage et Auer, 2006; Wilson, 2006).
Diagnostic radiographique
Une évaluation radiographique de la déviation angulaire est une étape indispensable pour déterminer le point de pivot et l’étiologie sous-jacente, mesurer l’angle de la déviation et évaluer la présence ou non d’une pathologie osseuse associée (Betsch, 2005; Trumble, 2005; Bramlage et Auer, 2006).
La prise des clichés radiographiques se fait avec un poulain debout, non tranquillisé ou à l’aide d’une tranquillisation mesurée pour éviter le relâchement musculaire. Il est utile de positionner le membre le plus naturellement possible avec des talons taillés symétriquement, en prenant appui sur ses quatre membres parallèles les uns aux autres pour éviter de créer artificiellement une déformation (Betsch, 2005; Witte, 2009).
L’interprétation des clichés radiographiques chez le poulain est délicate, car c’est facile de créer des images lésionnelles à partir d’images de croissance osseuse normale (Crevier-Denoix, 2005). Elle est réalisée sur deux parties: une analyse morphologique et une analyse géométrique.
L’analyse morphologique
L’analyse morphologique permet de relever différentes modifications radiographiques telles qu’un évasement de la physe, une plaque de croissance irrégulière, dissymétrique ou indistincte, élargie médialement ou latéralement ou épaisse dans les cas les plus avancés de déviation (Betsch, 2005). L’état et le degré d’ossification des os cuboïdes doivent être également évaluer (Wilson, 2006). Pour cela, la reconnaissance précoce de ces déformations est essentielle (García-López et Parente, 2011).
Les changements typiques observés avec l’écrasement du carpe et du tarse vont d’une incongruité subtile de la surface articulaire métacarpienne ou carpienne résultant d’un collapsus ou d’une hypoplasie de ces os (Figure 5-A), à une déformation évidente des os cuboïdes (fracture ou écrasement de ces os) (Figure 5-B) (Witte et Hunt, 2009).
Au niveau du tarse, l’os tarsal III situé au sommet de l’angle formé entre le tibia et le métatarse, subit de fortes pressions. Insuffisamment ossifié, son bord dorsal est comprimé, s’atrophie, voire se fracture (Figure 6). On parle alors de nécrose aseptique de l’os tarsal III (Witt et Hunt, 2009). Les poulains présentant une ossification incomplète des os du tarse et un affaissement de plus de 30% de l’os tarsal III et de l’os central ont un pronostic moins bon par rapport aux poulains avec moins de 30% d’effondrement, soulignant ainsi l’importance d’une reconnaissance et d’un traitement précoce (Ernst et al., 2020).
Un indice d’ossification squelettique SOI (Skeletal Ossification Index) est utilisé pour standardiser l’évaluation de la maturité osseuse chez les poulains nouveau-nés (Guillot et Lallemand, 2019). Il est établi à partir de deux clichés radiographiques, un dorso-palmaire/plantaire et l’autre latéro-médial, réalisés sur des poulains de moins de 2 semaines. L’indice se compose de quatre grades (Figure 7) (Adams et Poulos, 1988).
L’analyse géométrique
L’analyse géométrique a pour but de définir «le point de pivot» de la déviation. Pour ce faire, sur un cliché radiographique centré sur l’articulation d’intérêt, deux lignes droites coupant l’os long proximal et distal à la déviation en deux sont tracées (Baxter et Turner, 2002; Witte et al., 2004). L’angle aigu formé est l’angle de la déviation (Ernst et al., 2020). Le site de bissection des 2 lignes est défini comme le site primaire de la déviation et représente ce qu’on appelle le point de pivot. A partir de ce point, on distingue entre les localisations suivantes (Betsch, 2005; Bischofberger et Auer, 2019; KawCak, 2020):
• La physe ou juste proximalement, le site primaire de la déviation est alors la plaque de croissance métaphysaire (figure 8-A);
• La diaphyse d’un os long, lors d’incurvations diaphysaires associées (figure 8-B);
• L’épiphyse, le site primaire de la déviation est alors épiphysaire et/ou métaphysaire les deux étant la plupart du temps imbriqués;
• L’articulation ou distalement (cas du carpe), lors de laxités des tissus mous associées ou non au collapsus des os cuboïdes, ou dans le cas d’une anomalie des os du carpe ou des métacarpiens rudimentaires (Figure 8-C).
Plus la longueur d’os visible est importante, plus la mesure de l’angle sera précise. Au niveau du tarse, l’évaluation est plus délicate (le tibia n’est pas dans l’axe du canon). A cet effet, deux clichés de face (un dans l’axe de chaque diaphyse) et un cliché de profil (pour dépister un collapsus associé des os cuboïdes) sont souvent nécessaires (Greet et Curtis, 2003; Betsch, 2005; Smith 2010).
Gestion non chirurgicale des déviations angulaires
Le praticien dispose d’une panoplie de traitements pour la gestion des déviations angulaires d’une manière non chirurgicale. Le choix dépend de nombreux facteurs: âge du poulain, localisation et degré de la déviation, présence de rotation ou d’autres anomalies (Schambourg, 2004).
La plupart des déviations angulaires sont auto-correctrices et ne nécessitent qu’une prise en charge conservatrice. Elle implique généralement une combinaison de restriction d’exercice et de soins des pieds (parage ou extension du sabot) chez les poulains nouveau-nés atteints de déviations légères à modérées, et, dans certains cas, l’utilisation de bandages ou d’attelles (Auer et Rechenberg, 2006; Ruggles and McIlwraith, 2008; Ernst et al., 2020). Par contre, elle n’est pas recommandée comme première approche pour une déformation sévère de la physe chez un poulain de tout âge ou chez les poulains âgés (Ernst et al., 2020). L’approche conservatrice est également utilisée en complément d’une technique chirurgicale (Auer et Rechenberg, 2006).
Repos et exercice contrôlé
Des restrictions sur le plan d’exercice (combinée au parage correctif) sont utilisées de façon courante pour stimuler le développement musculaire et la mise sous tension des ligaments et des capsules articulaires, réduire le stress biomécanique sur le côté concave de l’angulation et assurer la modulation de la croissance physaire dans ses limites physiologiques (Betsch, 2005; Wilson, 2006). Ce mécanisme de compensation physiologique n’a lieu que si la plaque de croissance est non pathologique et encore fonctionnelle, c’est-à-dire au cours de la phase active de la croissance physaire. Par contre, la nature de cet exercice dépend du type, du degré de l’angulation et de l’étiologie sous-jacente (Greet, 2000).
Pour cela, il convient de contrôler radiographiquement l’évolution de la déviation et d’ajuster le traitement au fur et à mesure. Les moyens de cette rééducation passent par le confinement en stalle, la mise au box avec des sorties en main plusieurs fois par jour, le petit paddock (15x15m) ou la mise au pré quelques heures par jour seulement (Betsch, 2005).
Lors de déviation sévère ou complexe, l’exercice aggrave mécaniquement l’aplomb. Il est donc important de noter qu’il n’est toujours pas possible de prédire si le problème se résoudra avec le repos/exercice seul, et par conséquent, un temps précieux peut être perdu (Betsch, 2005; Bramlage et Auer, 2006). Cependant, dans beaucoup des cas, cette approche entraînera la correction de la déformation et permettra d’éviter les risques qui accompagnent une intervention chirurgicale parfois non nécessaire (Wilson, 2006).
Maréchalerie
La maréchalerie corrective a pour but de conserver une orientation et une forme physiologique soit par parage ou à l’aide d’extension, seule ou combinée. Le choix dépend de l’âge du poulain, de la sévérité et de la localisation de la déviation. Toutefois, il est important de rappeler que la modification des aplombs du membre distal par maréchalerie agit à tous les niveaux du membre, et peut créer éventuellement d’autres anomalies (Schambourg, 2004). En règle générale, jusqu’à l’âge de 4 mois, il convient de s’abstenir de corriger une déviation du carpe ou du tarse par une ferrure tant que les plaques de croissance du boulet sont encore actives et susceptibles de développer une déviation secondaire aux contraintes appliquées (la phase active de la croissance physaire est entre 0-100 jours au niveau du boulet) (Greet et Curtis, 2003; Schambourg, 2004).
Parage correctif
Le parage correctif (associé généralement au repos) est indispensable dans la gestion des défauts d’aplombs légers à modérés (Greet et Curtis, 2003; Betsch, 2005; Ernst et al., 2020). Il doit viser à produire un pied correct, c’est-à-dire assurer un équilibre latéro-médial et dorso-palmaire/ plantaire et réduire le stress inégal à travers le membre en parant le sabot du côté non suffisamment usé, puis d’arrondir la pince pour favoriser une bascule plus rapide (figure 9) (Greet, 2000; Betsch, 2005; Auer et Rechenberg, 2006; Smith, 2010).
Les directives du parage correctif pour les déformations angulaires des membres sont les suivantes (Greet et Curtis, 2003; Schambourg, 2004; Greet, 2015):
• Pour un varus du boulet, parer le pied de façon à ce que la sole soit perpendiculaire à l’axe des deux premières phalanges, c’est-à-dire parer la paroi interne.
• De même pour le valgus du boulet, le plan solaire du sabot doit être à 90° par rapport au grand axe des phalanges proximale et moyenne. Cela amène le membre à adopter une position verticale dans le plan axial. En pratique, il convient de parer la paroi externe.
• Pour le valgus du carpe (poulains de plus de quatre mois), le plan solaire du sabot doit être à 90° par rapport au grand axe du métacarpien III. Le pied doit être taillé pour avoir une forme symétrique. En pratique, il faut râper la paroi externe.
Il est recommandé de parer fréquemment et sans excès le pied avec une râpe (tous les 15 jours), pendant seulement quelques semaines à quelques mois, et de réaligner progressivement le plan solaire du pied pour rendre le pied plat (Schambourg, 2004; Betsch, 2005; Auer et Rechenberg, 2006). Un parage agressif et prolongé peut provoquer une surcorrection des déformations angulaires concomitantes, et dans les cas avancés une distorsion/torsion de la capsule du sabot sans correction de l’alignement des membres (Greet, 2000; Betsch, 2005; Ernst et al., 2020).
Extensions de sabot
Une autre forme de maréchalerie, qui utilise des extensions du sabot (Figure 10), consiste à étendre la surface solaire portante du pied dans le sens de l’alignement axial (Greet, 2000; Greet et Curtis, 2003; Greet, 2015).
Les extensions peuvent être utilisées à partir de 2 semaines d’âge chez le jeune poulain qui présente une angulation marquée à sévère ou s’il n’y a pas d’amélioration évidente dans les 2 semaines suivant le parage correctif. Par contre, il convient d’éviter de laisser les extensions du sabot sur place pour plus de 3 semaines ou de les utiliser comme seul traitement chez les poulains plus âgés, pour éviter le risque de développement des modifications dégénératives précoces du cartilage et des structures péri-articulaires et d’encastelure grave (Betsch, 2005; Auer et Rechenberg, 2006).
Attelles et plâtres
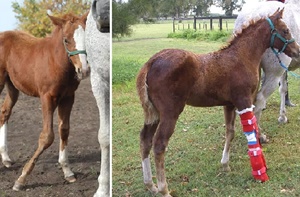
Les poulains présentant une hypoplasie des os du carpe et du tarse peuvent être traités efficacement avec des attelles ou des plâtres (Auer, 2006a). Le plâtrage doit être maintenu tant que l’ossification n’est pas atteinte, cela peut prendre de 2 à 4 semaines, et jusqu’à 2 mois selon le degré d’ossification au moment de la naissance (Betsch, 2005; Auer et Rechenberg, 2006). Ces dispositifs servent à maintenir le membre dans un bon alignement axial, permettant aux os hypoplasiques de s’ossifier correctement sans conséquences néfastes (Figure 12) (Auer et Rechenberg, 2006; Ernst et al., 2020). Ils sont également utilisés chez des poulains présentant une laxité périarticulaire sévère et des déformations angulaires graves, sous forme de bandages en association avec le repos en stabulation, pour maintenir le membre droit et permettre aux tissus périarticulaires et aux ligaments collatéraux de se renforcer (Jansson et Ducharme, 2005; Auer et Rechenberg, 2006; Witte et Hunt, 2009). Des attelles articulées spéciales sur mesure, qui permettent au poulain de déplacer les régions du carpe et du tarse tout en maintenant un alignement axial correct, ont été introduites sur le marché et peuvent être utilisées avec succès pour le traitement de ces déformations (Figure 11). Cependant, le traitement de la laxité périarticulaire par un support externe rigide a été débattu et contre-indiqué par certains auteurs (Auer et Rechenberg, 2006) et peut en effet retarder le renforcement des structures périarticulaires (Jansson et Ducharme, 2005; Auer et Rechenberg, 2006).
Selon la localisation de la déviation, le bandage ou le plâtre doit s’étendre à partir de la partie proximale du radius ou du tibia. L’incorporation du boulet et du pied dans la stabilisation doit être évitée afin de prévenir l’atrophie musculaire, l’hyperlaxité des structures tendineuses et l’exacerbation de l’affection (entraînant une chute du boulet) (Betsch, 2005; Jansson et Ducharme, 2005; Auer et Rechenberg, 2006; Witte et Hunt, 2009). L’attelle est changée tous les 3 à 4 jours et contrôlée quotidiennement pour éviter le risque de glissement. Le plâtre par contre doit être changé tous les 10 à 14 jours. En milieu hospitalier, il est recommandé de laisser l’attelle en place pendant 12 heures d’affilée pour prévenir les escarres, ou l’appliquer la nuit pendant que le poulain est moins actif (Jansson et Ducharme, 2005; Auer et Rechenberg, 2006). Des attelles de différents types de matériaux peuvent être utilisées et sont appliquées sur la face palmaire des membres antérieurs. L’utilisation du plâtre/attelle est plus difficile à appliquer aux membres postérieurs et même moins efficaces par rapport à son utilisation au niveau des membres antérieurs; cependant, dans les cas où il y a peu d’alternative, l’utilisation de ces appareils est indiquée (Bramlage et Auer, 2006).
L’évaluation radiographique est répétée à un intervalle de 2 semaines pour déterminer le degré d’ossification (figure 12) (Betsch, 2005). Le membre doit être bien rembourré avant l’application de l’attelle ou du plâtre pour prévenir les lésions cutanées de nécrose (surtout au niveau de l’emplacement de l’os carpien accessoire) en raison de la croissance rapide des jeunes poulains. En règle générale, le poulain doit se tenir correctement, sans aucun signe de chaleur, d’inappétence ou de douleur après l’application du support externe (Betsch, 2005; Auer et Rechenberg, 2006).
Thérapie des ondes de chocs extra-corporelles (Extracorporel shockwave therapy)
La thérapie des ondes de choc extracorporelles (ESWT) est une méthode non invasive utilisée pour retarder la croissance physaire du côté convexe du membre présentant une déviation angulaire (Figure 13) chez les poulains de plus de deux semaines (Bathe et al., 2006; Smith, 2010; Bussy et al., 2013; Greet, 2015; Koch et Goodrich, 2020). Elle a été suggérée comme alternative aux techniques chirurgicales pour le traitement des déviations angulaires chez les poulains (Bathe et al., 2006; Bussy, 2007).
Le mécanisme d’action exact par lesquels les ondes de choc affectent les os et les plaques de croissance n’a pas encore été déterminé. L’une des hypothèses est que les ondes de choc peuvent stimuler la néovascularisation et influencer de nombreux facteurs biologiques comme le FGF-2 et le TGF-β des tissus lésés (Bolt et al., 2004; Hausdorff et al., 2010).
Le protocole d’application des ondes de choc extracorporelles varie selon l’appareillage et les indications du fabricant. La dose usuelle est de 1500 à 2000 cycles à 15 Hz réglée entre 2,5 et 3 bars à intervalles hebdomadaires ou toutes les 2 semaines sur des poulains anesthésiés ou sous sédation pour un total de 1 à 6 séances (Bathe et al., 2006), et cela, jusqu’à ce qu’une correction clinique et radiographique soit obtenue (Smith, 2010; Bussy et al., 2013). Cette technique est utilisée en association avec des recommandations d’exercice et de maréchalerie qui dépendent du degré de la déviation angulaire (Bussy et al., 2013). Les premiers résultats rapportés sont encourageants, avec une plus grande efficacité sur les poulains atteints des déformations légères et modérées, en particulier du carpe et du tarse (Greet, 2015).
La bande de kinésiologie
La bande de kinésiologie est une technique non invasive et peu couteuse, qui utilise des bandes adhésives élastiques. Elle est couramment utilisée pour réduire la formation d’œdèmes et les gonflements généralisés, améliorer le flux sanguin et favoriser la stabilisation des structures anatomiques. Bien que le mécanisme d’action exacte et son effet sur les structures musculo-tendineuses ne sont pas encore bien élucidés, avec un débat important sur son efficacité clinique, des effets bénéfiques ont été reportés par des vétérinaires équins et des propriétaires de chevaux (Mikail et al., 2019; Koch et Goodrich, 2020).
Son utilisation dans le traitement de «l’hallux valgus» en médecine humaine a montré une amélioration significative de la douleur et de l’alignement articulaire après 10 jours de mise en œuvre (Karabicak et al., 2015). De son tour, son utilisation chez une pouliche de 30 jours et un poulain de 60 jours avec un valgus carpien droit de 153° et un valgus carpien gauche de 162° respectivement, a montré une amélioration significative après 15 jours du traitement (180° pour la pouliche et 177° pour le poulain). Dans les deux cas, un «VetkinTape» de 20 cm a été appliqué avec un étirement de 30 % sur la face médiale du membre affecté pour soutenir le ligament collatéral médial du carpe et un «VetkinTape» de 10 cm a été enroulé sur les deux extrémités du premier ruban pour ancrer cette première bande (figure 14) (Mikail et al., 2019). L’adaptation de cette technique pour le traitement des déviations angulaires en médecine équine est nouvelle et manque de preuves scientifiques sur son efficacité, mais elle montre un potentiel prometteur pour les déformations angulaires chez les poulains.
DISCUSSION
Le traitement conservateur commence tout d’abord par la gestion de l’alimentation du poulain. Certains auteurs recommandent de sevrer précocement le poulain ou de diminuer sa ration alimentaire surtout chez les poulains à croissance rapide (Schambourg, 2004). Pour pouvoir choisir entre les différents traitements conservateurs disponibles, Il convient de les aborder par étiologie.
La laxité des tissus mous périarticulaires
Chez les poulains nouveau-nés, la laxité périarticulaire est la cause la plus probable des déviations angulaires (Wilson, 2006). La croissance résout généralement les déformations légères à modérées dans les 2 à 4 premières semaines de vie (Jansson et Ducharme, 2005). D’après Wilson (2006), le clinicien qui souhaite instaurer un traitement pourra recommander de garder le poulain en stalle pour une courte période. Par contre, Jansson et Ducharme (2005) pensent que la restriction d’exercice est contre-indiquée, car l’exercice est important pour le développement musculaire normal et le renforcement des tissus mous impliqués, donc à la résolution de la déviation. Par conséquent, ils ont conseillé de placer la jument et le poulain dans un petit enclos (15 × 15 m) ou de les garder dans une grande stalle et autoriser l’accès fréquemment à un petit enclos pendant quelques heures pour éviter une fatigue excessive. Auer et Rechenberg (2006) suggèrent un programme d’exercice différent qui consiste en une marche quotidienne de 10 à 20 minutes avec la jument si l’ossification des os cuboïdes est normale ou 3 à 5 minutes de natation par jour si les installations sont disponibles.
Le traitement de la laxité périarticulaire par coaptation externe est controversé. En général, la coaptation externe est indiquée seulement en cas d’hypoplasie des os cuboïdes d’après Betsch (2005) et ne doit pas être utilisée pour la laxité périarticulaire (avec ossification osseuse cuboïde normale), car elle aggraverait le problème (Bischofberger et Auer, 2018). Cependant, dans les cas de laxités sévères (par exemple, dans le cas où les os carpiens du poulain frottent les uns contre les autres (Wilson, 2006)), il est peu probable d’avoir une correction spontanée. Dans ce cas, un support mécanique externe sous la forme d’un bandage rigide ou d’une attelle ou d’un tube plâtré peut être appliqué pour redresser le membre en plus du repos en stalle, et permettre aux tissus périarticulaires et aux ligaments collatéraux de se renforcer en 2 à 4 semaines (Jansson et Ducharme, 2005).
Ossification incomplète des os cuboïdes
Les déformations angulaires du membre qui résultent d’une ossification incomplète avec des membres droits doivent être traitées avec le repos et la restriction d’exercice jusqu’à ce qu’une ossification complète se produise (Bischofberger et Auer, 2018). La restriction de l’activité physique doit être réduite au minimum car le développement normal du poulain en tant que potentiel athlète dépend de l’exercice (Bischofberger et Auer, 2018). La période moyenne est de 14 à 21 jours (Wilson, 2006). Lemonnier et Dugardin (2005) préconisent une période de 2 semaines qui s’avère habituellement suffisante pour obtenir le résultat souhaité. Alors que Auer et Rechenberg (2006) conseillent de maintenir le repos en stabulation pendant un maximum d’1 mois. Pendant cette période, les radiographies doivent être répétées à des intervalles de 2 semaines pour évaluer la progression de l’ossification (Auer et Rechenberg, 2006; Bischofberger et Auer, 2018).
Les poulains avec une ossification incomplète combinée à des membres qui ne sont pas droits ou avec une ossification incomplète des os cuboïdes du tarse ou avec un mauvais alignement des articulations peuvent nécessiter une coaptation externe pour assurer l’alignement axial de l’articulation et permettre la maturation des os cuboïdes, au point qu’ils ne sont pas soumis à un poids dépassant leur capacité (Auer et Rechenberg, 2006; Bramlage et Auer, 2006). Dans de rares cas (c.-à-d. poulains jumeaux), il peut être nécessaire de poursuivre la pose des attelles jusqu’à ce que le poulain atteigne l’âge de 2 mois. Cependant, il est important de garder le membre bandé pendant 4 à 5 jours supplémentaires après le retrait de l’attelle ou du plâtre et l’exercice doit être repris progressivement (Bischofberger et Auer, 2018). Trumble (2005) préfère de laisser le membre libre pour une stimulation des tissus mous. En effet, Lemonnier et Dugardin (2005) ont remarqué à plusieurs reprises, une hyperlaxité ligamentaire associée à une descente du boulet (“bas jointé”) au retrait du bandage.
Croissance asynchrone des plaques de croissance
Les déformations angulaires acquises des membres résultent d’une croissance osseuse longitudinale disproportionnée au niveau de la physe (Witte et Hunt, 2009). Les déformations angulaires légères à modérées de ce type répondent favorablement à un traitement conservateur basé sur le confinement en stalle et le parage correctif des sabots et, éventuellement, l’application de fers à coller (Parente, 2003), qui doit toujours être considéré comme le traitement initial si possible (Ernst et al., 2020). Si l’angulation est inférieure à 10°, le repos en stalle et l’exercice contrôlé (limité, s’il y au petit paddock) peut être poursuivi pendant 4 à 6 semaines (Greet, 2000; Auer et Rechenberg, 2006). Si les radiographies démontrent que la déformation n’est pas d’origine articulaire, l’application d’attelles et de plâtres est inefficace et, en fait, contre-indiquée, car il en résulte presque invariablement des escarres cutanées sans correction de la déformation (Jansson et Ducharme, 2005; Bischofberger et Auer, 2018).
L’utilisation de la thérapie des ondes de choc extracorporel (ESWT) chez des poulains au niveau de la partie convexe de la plaque de croissance a montré des résultats prometteurs pour le traitement des déviations d’origine physaire. Bathe et al. (2006) ont eu un taux de réussite de 88 % (15 poulains sur 17), qui a été revendiqué dans les 15 à 76 jours. Une autre étude menée par Bathe et Hilton (2008) a révélé que 57/67 (85 %) poulains ont subi un ESWT réussi entre 2004 et 2007. L’ESWT a été également utilisé pour le traitement poulains présentant un valgus carpien de degré variable (de léger (5° à 8.9°), modéré (9° à 11.9°) à sévère (12° à 21.6°). Le succès du traitement, défini comme un angle de déformation en valgus inférieur à 5˚, a été atteint chez tous les poulains (n=64), à l’exception de 5 poulains lors de la dernière évaluation radiographique (Bussy et al., 2013). L’impression clinique est que les poulains présentant des déformations légères traitées avec un parage de sabot ou des extensions de sabot seules ont besoin de plus de temps pour la résolution des déformations angulaires des membres que ceux traités avec ESWT (Bussy et al., 2013).
CONCLUSION
Les déviations angulaires chez le poulain ne sont pas forcément définitives s’ils sont diagnostiqués et traités à temps, puisqu’avant la fin de la croissance, la conformation peut encore se modifier.
La panoplie de choix pour la prise en charge de ces déviations prête à confusion. Cependant, les méthodes conservatrices permettent une correction spontanée dans les déviations légères à modérées. Elles peuvent associer une période de repos, un parage des pieds, une ferrure corrective avec extension ainsi que l’utilisation des ondes de chocs extracorporelles.
Enfin, pour les déviations sévères ou qui ne répondant pas favorablement à la méthode conservatrice, la chirurgie est indiquée.
Au bout de cette synthèse bibliographique, il est possible de tirer une conclusion majeure : le diagnostic et la prise en charge précoce sont d’une grande importance et présentent un effet considérable sur les chances de résolution complète. Plus le poulain est âgé et en fin de croissance, plus le risque d’avoir des lésions irréversibles augmentent, et les techniques de correction seront complexes et invasives. Par contre, si le vétérinaire est impliqué dans l’évaluation du poulain à un stade précoce, des traitements conservateurs peuvent être essayés en premier et la fenêtre d’opportunité de leur choix serait beaucoup plus large.
Bibliographie
Adams R., Poulos P. A. (1988). Skeletal ossification index for neonatal foals. Veterinary Radiology, 29: 217-222.
Anonyme 1. www.redboot.com.ar/diapositivas (28/11/2021).
Auer J.A. (1999). Angular limb deformities, In Equine Surgery, Eds: Auer J.A. & Stick J.A., Philadelphia, WB Saunders, 736–752.
Auer J.A. (2006). Angular limb deformities, In Equine Surgery (3rd Edition), Eds: Auer J.A. & Stick J.A., St. Louis, Mo, Saunders Elsevier, 1130-1149.
Auer J.A., Rechenberg B. (2006). Treatment of Angular Limb Deformities in Foals. Clinical Techniques. Equine Practice, 5: 270-281.
Bathe A.P., Hilton R.L. (2008). Treatment of angular limb deformities using radial extracorporeal shock wave therapy. Proceedings of the 14th Congress of the European Society of Veterinary Orthopaedics and Traumatology. Munich, 222-223.
Bathe A.P., Rowlands D.S., Boening K.J. (2006). Treatment of Angular Limb Deformities Using Radial Extracorporeal Therapy: A Prospective Clinical Trial,” Proceedings of the 9th International Congress of World Equine Veterinary Association, Marrakech, 22-26 January, 167-168.
Baxter G.M., Turner A.S. (2002). Diseases of the bone and related structures. In Adams’ Lameness in horses (7th Edition) Ed: Stashak T.S., Lippincott Williams and Wilkins, Philadelphia, 401-457.
Betsch J. (2005). Reconnaissance, évaluation et gestion des déviations angulaires du poulain. Pratique vétérinaire équine, 37: 45-58.
Bischofberger A.S., Auer A.J. (2019). Angular Limb Deformities, Equine Surgery (5th Edition), Eds: Auer J.A., Stick J.A., Kümmerle J.M. & Prange T., W.B. Saunders, Chap 87: 1471-1490.
Bolt D., Burba D., Hubert J. (2004). Determination of Functional Changes in Palmar Nerves after Nonfocused ESWT in Horses. American Journal of Veterinary Research, 65: 1714-1718.
Bramlage L.R., Auer J.A. (2006). Diagnosis, assessment and treatment strategies for angular limb deformities in the foal. Clin. Tech. Equine Pract., 5: 259-269.
Bussy C. (2007). Déviations Angulaires et Ondes de Choc. Le Point Vétérinaire, 275: 39-43.
Bussy C., Auzas F., Muñoz J.A. (2013). Clinical Use of Extracorporeal Shockwave Therapy (ESWT) for the Treatment of Carpus Valgus Deformities in Young Foals: A Retrospective Study of 64 Cases (2006-2009). Open Journal of Veterinary Medicine, 3: 46-51.
Crevier-Denoix N. (2005). Radiologie du squelette I: régions distales des membres et membre antérieur. Pratique vétérinaire équine, 37: 15-25.
Dubois B.B., Rodgerson D.H. (2020). Single-incision percutaneous drilling technique to achieve hemiepiphysiodesis of the distal metacarpus in foals with metacarpophalangeal varus deformities. Veterinary Surgery, 49: 940-946.
Ernst N. S., Trumble T. N., Baxter G. M. (2020). Flexural Limb Deformities, Chapter 10- Lameness in the Young Horse, In Adams & Stashak Lameness in Horses, Seventh Edition, Edited by Gary M. Baxter, John Wiley & Sons, 1059-1070.
García-López J.M., Parente E.J. (2011). Angular Limb Deformities (Chap. 58), In Diagnosis and Management of Lameness in the Horse (2nd Edition), Ross M.K., Dyson S.J., W.B. Saunders (Eds), 640-645.
Greet T. (2000). Managing flexural and angular limb deformities: the Newmarket perspective. Proc. Am. Assoc. Equine Pract., 46: 130–136.
Greet T. (2015). Angular and flexural limb deformities in foals and yearlings. Veterinary Nursing Journal, 30: 352-355.
Greet, T.R.C., Curtis S.J. (2003). Foot management in the foal and weanling. Vet. Clin. Equine, 19: 501–517.
Guillot M., Lallemand E. (2019). Déviations angulaires chez le poulain: présentation des affections et modalités diagnostiques. Pratique Vétérinaire Équine, 203: 6-13.
Hausdorf J., Sievers B., Schmitt-Sody M., Jansson V., Maier M., Mayer-Wagner S. (2010). Stimulation of Bone Growth Factor Synthesis in Human Osteoblasts and Fibroblasts after Extracorporeal Shock Wave Application. Archive of Ortho. Traum. Surg., 24: 10-18.
Jansson N., Ducharme N.G. (2005). Angular Limb Deformities in Foals: Treatment and Prognosis. Compendium, 134-146.
Karabicak G.O., Bek N., Tiftikci U. (2015). Short-Term Effects of Kinesiotaping on Pain and Joint Alignment in Conservative Treatment of Hallux Valgus. Journal of manipulative and physiological therapeutics, 38: 565-571.
KawCak C. (2020). The Carpus. In Adams and Stashak’s lameness in horses (7th Edition), Lameness of the Proximal Limb (Chapter 5). Ed: Baxter G.M., Wiley-Blackwell. 597-686.
Koch D.W., Goodrich L.R. (2020). Principles of Therapy for Lameness: Topical/Local. In: Adams and Stashak’s Lameness in horses. 881-885.
Lemonnier J.P., Dugardin D. (2005). Incidence défavorable de la prématurité sur l’articulation tarsienne du poulain. Pratique Vétérinaire Équine, 37: 33-40.
Mikail S., Araujo M.V.I., Jatobá M.V., Silva M.I.G., Carvalho L.M. (2019). Correction of angular limb deformities in foals using kinesiology taping. Acta Veterinaria Scandinavica, 61: 6.
Morisset S. (2005). L’étiopathogénie des déviations angulaires et sagittales chez le poulain. Pratique Vétérinaire Equine, 37: 35-43.
O’Grady S.E. (2017). Routine trimming and therapeutic farriery in foals. Vet. Clin. North Amer. Equine Pract., 32: 267–288.
Parente E.J. (2003). Angular limb deformities, in Ross MW, Dyson SJ (eds): Lameness in the Horse. Philadelphia, 557–561.
Redden R.F. (2011). How to Evaluate Foot Flight and Leg Alignment. AAEP Proceedings, 57: 407-414.
Robert C., Valette J.P., Denoix J.M. (2013). Longitudinal development of equine forelimb conformation from birth to weaning in three different horse breeds. Vet. J., 198: 75-80.
Ruggles A.J., McIlwraith C.W. (2008). Management of angular and flexural disorders in foals. Proc AAEP Focus Meeting: First Year Life, Austin, 60–67.
Schambourg M. (2004). Comment traiter les déviations angulaires chez le poulain. Le nouveau praticien vétérinaire équine, 1:44-47.
Smith L. (2010). Treatment of angular limb deformities in foals. In Practice, 32: 156-162.
Smith L. (2015). Angular limb deformities in foals: a clinical review. Livestock, 20: 111-117.
Trumble T.N. (2005). Orthopedic disorders in neonatal foals. Vet. Clin. North Am. Equine Pract., 21: 357–385.
Watson R. (2016). An overview of limb deformities in foals. Equine Health, 29:30-33.
Wilson D.G. (2006). Les déformations angulaires des membres chez le poulain: prise en charge et conséquences de l’angulation sur la performance athlétique. La médecine vétérinaire des Grands Animaux: Rondes cliniques, 6(1).
Witte S. (2009). A review of angular limb deformities. Equine Vet. Edu., 21: 378–387.
Witte S., Hunt R. (2009). A review of angular limb deformities. Equine Vet. Educ., 21: 378–387.
Witte S., Thorpe P.E., Hunt R.J., Spirito M.A., Rodgerson D.H. (2004). A lag-screw technique for bridging of the medial aspect of the distal tibial physis in horses. J. Am. vet. med. Ass., 225: 1581-1583.
Publié-e
Numéro
Rubrique
Licence

Revue Marocaine des Sciences Agronomiques et Vétérinaires est mis à disposition selon les termes de la licence Creative Commons Attribution - Pas d’Utilisation Commerciale - Partage dans les Mêmes Conditions 4.0 International.
Fondé(e) sur une œuvre à www.agrimaroc.org.
Les autorisations au-delà du champ de cette licence peuvent être obtenues à www.agrimaroc.org.

